V. Evolutionsbelege
In den vorangegangenen Kapiteln haben wir die gängigsten, von Evolutionsgegnern erörterten Themenbereiche behandelt und dabei verschiedentlich schon in groben Zügen umrissen, welche Beobachtungen für (transspezifische) Evolution sprechen. Wir haben ferner festgestellt, daß wir weder für Evolution noch für sonst ein in der Naturwissenschaft theoretisiertes Faktum Beweise im streng logischen Sinne, sondern immer nur Belege anführen können, welche in unserem Falle die Abstammungshypothese mit mehr oder minder hohem Grade der Sicherheit abstützen. Der Übersichtlichkeit halber wollen wir die wichtigsten Belege und Gegenargumente in diesem Kapitel zusammenfassend besprechen.
1. Die abgestuften Ähnlichkeiten in der vergleichenden Biologie
Seit DARWINs epochalem Werk "On the Origin of Spezies" von 1859 zählen die Ergebnisse der vergleichenden Anatomie sicher zu den nachhaltigsten Belegen für die Abstammungs- oder Deszendenzhypothese. Streng genommen liegen bereits seit LINNE Befunde vor, in deren Licht die Evolutionsvorstellung nicht mehr nur als vage Spekulation, sondern als begründete, wohlbestätigte Annahme erscheint (JAHN et al., 1982, S. 256 f.). Doch erst im Rahmen des von DARWIN vorgeschlagenen, theoretischen Evolutionsmechanismus (Variation und natürliche Zuchtwahl) ergeben sich eine Reihe logischer Folgerungen (Deduktionen) aus der Abstammungshypothese, die mit der Beobachtung im Einklang stehen:
Daß strukturelle "Information" genetisch an die Nachkommen vererbt wird, ist einleutend. Die Vererbung stellt, salopp formuliert, ein "Gedächtnis" dar, "einen Speicher für alle Erfolge, die das Leben jemals errungen hat." (v. DITFURTH, 1987, S. 40). Wenn also die evolvierten Merkmale von Generation zu Generation weitervererbt werden, wenn zugleich aber auch ein allmählicher Wandel der Arten stattfindet und wenn die Abstammungshypothese stimmt, dann ergibt sich die logische Folgerung, daß zwischen den Arten eine (abgestufte) Formenähnlichkeit bestehen muß, die sich vom morphologischen bis hinab zum molekularen Bereich erstreckt (v. DONGEN und VOSSEN, 1984).
So ist für uns die Abstammung offensichtlich, wenn ein Kind seinen Eltern "wie aus dem Gesicht geschnitten" oder der Enkel in seinem Verhalten "ganz der Opa" ist. Entsprechend kann auch an der Verwandtschaft zwischen den verschiedenen Arten kein rational begründeter Zweifel bestehen, wenn man hier auf tiefgreifende Formenähnlichkeiten stößt - vorausgesetzt, man erkennt die Standards wissenschaftlichen Argumentierens an.
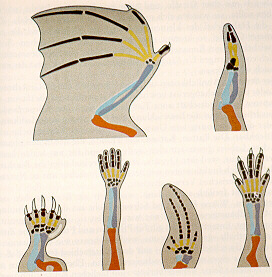
Vergleicht man die Organismen verschiedener Arten und Gruppen miteinander, stellt man tatsächlich fest, daß zwischen ihnen mehr oder minder große Ähnlichkeiten existieren (einen guten Überblick über die Evolutionsbelege verschafft OSCHE, 1979, S. 11-30). Als klassisches Beispiel seien die Extremitäten der Vierbeiner (Tetrapoden) genannt, die bei den Arten zum Teil völlig unterschiedliche Funktionen übernehmen und dennoch einen fast identischen Feinbau besitzen. Vögel, Säugetiere, Reptilien - sie verfügen gleichermaßen über Oberarmknochen, Elle, Speiche, Handwurzelknochen, Mittelhand, sowie über (zumeist fünf) Finger. Auch die Extremitäten der Wale gleichen denjenigen von Vögeln und Primaten (vgl. Abbildung 1). REMANE et al. weisen darauf hin, daß gerade solch komplexe Merkmalsgruppierungen, die bei verschiedenen Arten detailgetreu in Erscheinung treten, aber im Dienste verschiedener Funktionen stehen, auf Abstammung von einem Vorfahren hindeuten (REMANE et al., 1973, S. 43; ähnlich OSCHE, 1979, S. 12; GOULD, 1987, S. 25; BERCK, 2002, S. 35 ff.; NEUKAMM, 2002, S. 40f.). Wie eine solche Datensituation völlig ohne Vererbung und transspezifische Evolution erklärt werden könnte, ist meines Erachtens bislang noch nicht überzeugend aufgezeigt worden.
Die strukturellen Übereinstimmungen lassen sich in neuerer Zeit bis in den molekularen Bereich hinein verfolgen, wobei sich zeigt, daß (fast) alle rezenten Lebewesen nicht nur denselben genetischen Code besitzen, sondern auch in weiten Teilen über ein nahezu identisches Repertoire an Genen, Biomolekülen und Stoffwechselprozessen verfügen. Selbst Verhaltensweise und Mimikri sind bei höheren Säugetieren ausgesprochen ähnlich. Die vergleichende Verhaltensforschung hat sich daher als moderner Zweig in der Evolutionsforschung etabliert und versucht, Verhaltensmerkmale zur Stammbaumerstellung heranzuziehen (WINKLER, 1994).
Abbildung 1:
Ähnlichkeiten im Skelettbau der Extremitäten von Vierfüßern (Tetrapoden). Von oben nach unten und von links nach rechts: Fledermaus, Vogel, Maulwurf, Mensch, Wal und Eidechse. Trotz verschiedener funktioneller Beanspruchung ist der Feinbau des Skeletts bei den Tetrapoden praktisch identisch.
Wenn sich durch Aufspaltung und Umwandlung von Stammarten schrittweise neue Arten gebildet haben und einmal getroffene "Entscheidungen", die sich als erfolgreich herausgestellt haben, kaum mehr rückgängig zu machen oder bestenfalls nur sehr eingeschränkt veränderbar sind, müssen sich die vererbten Merkmale (Homologien) auch in eine abgestufte Rangfolge bringen lassen (AX, 1984; WÄGELE, 2000). Daraus resultiert dann im Idealfall eine gesetzmäßige Beziehung zwischen den Merkmalen oder anders ausgedrückt: eine abgestufte Formenähnlichkeit, ein hierarchisches System konsequent ineinandergeschachtelter Organismenklassen (MAHNER und BUNGE, 2000, S. 230; Kapitel II).
Auch diese logische Erwartung findet ihre (allerdings nicht vollkommene) Entsprechung in der Wirklichkeit. Man findet die ähnlichen Merkmale bei den verschiedenen Arten nicht völlig regellos kombiniert, sondern häufig gesetzmäßig miteinander verbunden vor:
Aus der abgestuften Merkmalsbeziehung läßt sich mit anderen Worten der Schluß ziehen, daß die Formenmannigfaltigkeit das Ergebnis einer stammesgeschichtlichen Entwicklung ist, die sich in langen erdgeschichtlichen Zeiträumen vollzogen hat.
Diese These wird auch durch den Fossilienbefund gestützt, der ebenfalls zu den klassischen Belegen für eine gemeinsame Stammesgeschichte der Arten zählt. Weil sich in der Evolution die schrittweise Abänderung von Merkmalen vollzieht, ergibt sich nämlich die Forderung, daß es fossil erhaltene Arten geben muß, die in ihren mosaikartigen Merkmalsgefügen "zwischen" den heute existierenden Organismengruppen stehen (1). Desweiteren muß "diese Verknüpfung zeitlich geordnet sein, sich also auch in der geologischen Zeit schrittweise dem Typus angliedern" (REMANE et al., 1973, S. 26).
Betrachten wir die fossil überlieferten Tier- und Pflanzenarten, so stellt man - ungeachtet der Lückenhaftigkeit der Dokumentation - in der geochronologischen Abfolge der Fossilien tatsächlich keinen chaotischen, sondern einen geordneten Formenwandel fest. Die Lebewesen haben sich im Laufe der Zeit in ihrem Bau immer mehr den heutigen Formen angeglichen. Zu den Paradebeispielen zählt beispielsweise die Formenreihe in der Pferdeevolution (OSCHE, 1979, S. 22). Auch die Entstehung der Wale und Rüsseltiere ist fossil relativ gut und durch zahlreiche Übergangsformen abgestützt (siehe Abbildung 2).
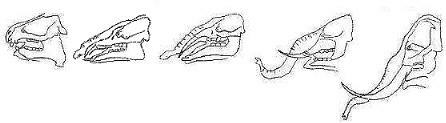
Abbildung 2: Die Entstehung von Rüssel und Stoßzähnen bei den Rüsseltieren. Von links nach rechts: Moeritherium (Eozän), Palaeomastodon (Oligozän), Mastodon (Miozän), Stegomastodon (Pliozän), Mammonteus (Pleistozän). Nach THENIUS und HOFER, 1960.
Ein weiteres Beispiel für evolutionäre Umwandlungen findet man in der Entwicklung des sekundären Kiefergelenks der Säugetiere aus dem primären Kiefergelenk der "Reptilien". Fast alle nennenswerten Zwischenstadien liegen paläohistorisch dokumentiert vor, einschließlich solcher Formen, bei denen beide Kiefergelenke funktional nebeneinander vorliegen (beispielsweise bei Diarthrognathus).
Zusammenfassend läßt sich also festhalten, daß die deduktiven Folgerungen der DARWINschen Abstammungslehre relativ gut mit der Beobachtung in Einklang stehen. Sie führt die lange Zeit disparat nebeneinanderstehenden Erscheinungen zusammen und erklärt sie völlig zwanglos.
Die hypothetico-deduktive Methode der "Beweisführung" wird von Evolutionsgegnern jedoch stark kritisiert. JUNKER und SCHERER betonen die "Theoriebeladenheit" der Methode und zeigen, daß abgestufte Formenähnlichkeiten nicht als streng logische Beweise, sondern bestenfalls als fehlbare Indizien ("Deutungen") für Evolution verstanden werden dürfen (JUNKER und SCHERER, 1998, S. 155).
Außerdem wird darauf hingewiesen, daß die Bestimmung von abstammungsbedingten Ähnlichkeiten (Homologien) erheblich dadurch erschwert wird, daß sich die Merkmale in aller Regel nicht in eine vollkommene Hierarchie einfügen lassen. Einfacher ausgedrückt: Viele Ähnlichkeitsmerkmale gehen nicht auf gemeinsame Abstammung zurück (Homologie), sondern sind unabhängig voneinander entstanden (Konvergenz, Analogie). Konvergente Merkmale (wie etwa die Flossen von Wal und Hai) werden in der klassischen Evolutionstheorie als das Ergebnis der Anpassung an gleichartige Umweltbedingungen verstanden (OSCHE, 1979, S. 53-56). Eine Entscheidung, welche Merkmale Homologien verkörpern und welche nicht, läßt sich oft nur durch theoretische Überlegungen und Merkmalsgewichtungen entscheiden, nicht aber durch die "nackten", "theoriefreien Beobachtungstatsachen". Dazu JUNKER:
"Die (homologe oder analoge) Ähnlichkeit der Organismen an sich gibt über ihre Ursache keine eindeutige und zwingende Auskunft. Daher liefern Ähnlichkeiten keinen Beweis für Evolution (...) Wenn nach bestimmten Kriterien erkannte Homologien sich auch als Konvergenzen herausstellen können (...) ist die übliche Schlußfolgerung vom Vorliegen einer Homologie auf gemeinsame Abstammung nicht mehr möglich."
(JUNKER, 2002, S. 28, 40)
Schließlich wird betont, daß man für Ähnlichkeiten auch funktionale Gründe finden könne, die einen Schöpfer dazu "bewogen" haben könnten, verschiedene Arten mit ähnlichen Strukturen auszustatten, sofern sie ähnliche Funktionen zu erfüllen haben. Daher halte die prinzipielle Unmöglichkeit, Ähnlichkeiten sicher auf die historische Kontingenz gemeinsamer Abstammung zurückzuführen (und Funktionsähnlichkeit als mögliche Ursache zweifelsfrei auszuschließen) der Schöpfungsalternative einen Weg offen (JUNKER, 2002 b).
Man muß es zugeben: Die Argumentation ist logisch unangreifbar. Dennoch läßt sich das gewünschte Ziel, nämlich die Schöpfungsthese wissenschaftlich zu rechtfertigen, nicht erreichen, weil der Gedankengang methodologisch fragwürdig ist. Folgende Einwände lassen sich anführen:
Wenn behauptet wird, daß die Übereinstimmung zwischen einer theoretischen Erwartung und einer Beobachtung die Theorie nicht beweise, so ist das streng logisch gesehen zwar korrekt. Die Behauptung enthält aber kein wissenschaftliches Argument gegen die Theorie, weil praktisch alle Fakten (seien es historische oder gegenwärtige) auf diese Weise erschlossen werden (vgl. Kapitel Ib.2.1). Man kann dann nur noch die hypothetisch-schlußfolgernde Methode der Naturwissenschaft generell infragestellen und damit den Boden der wissenschaftlichen Argumentation verlassen.
Die Schöpfungshypothese ist keine wissenschaftliche Alternative zur Evolutionstheorie, weil Wissenschaft vorrangig nach Erklärungen für bislang unverstandene Phänomene sucht. Solche Erklärungen, wie wir sie im Lichte der Abstammungshypothese vorgestellt haben, kann man mit der Schöpfungshypothese aber nicht vornehmen. Denn was der "Kreator" erschaffen hat, wie und warum er das alles so und nicht anders erschuf, wie er es getan hat, kann man mit den "natürlichen" Mitteln, die uns zur Verfügung stehen, prinzipiell nie erforschen. Der Schöpfer braucht sich weder zu erkennen geben, noch muß er sich innerweltlichen Gesetzen beugen oder mit Materie wechselwirken. Der Ratschluß des "intelligenten Designers" bleibt von vorn herein ein unlösbares Mysterium und erklärt daher nichts. (2)
Auch das "Homologieproblem" ist kein stichhaltiges Argument gegen die Abstammungshypothese. Selbst wenn man vielfach nicht sicher entscheiden kann, bei welchen Ähnlichkeitsmerkmalen es sich um abstammungsbedingte Homologien handelt, bedeutet das nicht, daß es keine Homologien und keine Evolution gibt. (3)
2. Höherentwicklung (Anagenese) und Entropie
Wenn es eine transspezifische Evolution gibt, muß eine Anagenese (Höherentwicklung) - im Sinne des schrittweisen Auftretens immer komplexerer Lebensformen - im Fossilienbefund festzustellen sein. Diese Folgerung leuchtet, wie v. DITFURTH betont, unmittelbar ein:
"Bevor man sich den Luxus der Anschaffung von Flügeln, Antennen oder anderen Spezialausrüstungen leisten kann, müssen erst einmal die elementaren, lebenserhaltenden Funktionen, etwa die des Stoffwechsels, gewährleistet sein. Die 'Generalien' des Lebens mußten früher verwirklicht werden als alle über das Notwendigste hinausgehenden speziellen Ausstattungen."
(v. DITFURTH, 1987, S. 41)
Diese Erwartung läßt sich paläontologisch bestätigen. Betrachtet man den Fossilienbefund, treten im Lauf der geologischen Zeit immer komplexere Lebewesen in den Schichten in Erscheinung. In stark vereinfachter Form läßt sich paläontologisch ungefähr folgende Anagenese nachzeichnen:
erste Metazoen mit Ganglien (Nesseltiere) -> Metazoen mit Strickleiternervensystem (Anneliden, Mollusken,...) ->
Chordata mit dorsalem Zentralnervensystem (Manteltiere) -> erste Wirbeltiere mit Hirnstamm (Fische) ->
erste Wirbeltiere mit R-Komplex (Reptilien) -> erste Wirbeltiere mit limbischem System (Vögel) /
erste Wirbeltiere mit Neocortex (Säugetiere) -> Primaten mit hochentwickeltem Neocortex (Mensch) -> ???
Eine Höherentwicklung von einfach zu komplex organisierten Strukturen hat also nicht nur während der kosmischen und chemischen Evolution stattgefunden, sondern auch während der Geschichte des Lebens. Entsprechend läßt sich die "Reihenfolge der Typen" nicht nur paläontologisch, sondern, wie obiges Schema andeutet, auch vergleichend anatomisch rekonstruieren. Ordnet man etwa die einzelnen Gruppen der Wirbeltiere hinsichtlich der Komplexität des Gehirns, so entsteht eine Reihe "Fische - Amphibien - "Reptilien" - Vögel/Säugetiere". Die Evolutionsreihe ist praktisch identisch mit der zeitlichen Aufeinanderfolge der fossilen "Typen" in der geologischen Schichtung, die Abstammungslehre stellt mit anderen Worten die plausibelste Erklärung für diese isoliert betrachtet zunächst rätselhaft erscheinenden Beobachtungen (ähnlich BAUER, 1981, S. 331).
Evolutionsgegner sehen das naturgemäß anders. So behaupten beispielsweise JUNKER und SCHERER, aus dem bloßen Formenvergleich sei eine Höherentwicklung nicht ableitbar. Die These, daß Höherentwicklung immer nur in die Merkmale "hineingelesen" werden können, erklären die Autoren am Beispiel des Herz-Blutkreislaufsystems der Wirbeltiere. Da ein mit dem Herzkreislaufsystem eines Säugetieres ausgestatteter Fisch ökonomisch benachteiligt sei, benötige er es nicht. Deshalb könne beim Vergleich der Herz-Blutkreislaufsysteme nicht auf eine Höherentwicklung vom Fisch zum Säuger geschlossen werden (JUNKER und SCHERER, 1998, S. 157).
Die Kritik ist berechtigt, wenn man mit dem Begriff "Höherentwicklung" (wie das leider heute noch allzu oft geschieht) eine Wertung vornehmen wollte, beispielsweise im Sinne einer fortlaufenden Vervollkommnung, besseren Anpassung oder Ökonomisierung. Wenn man Höherentwicklung allerdings als rein beschreibenden Begriff gebraucht und beispielsweise die Komplexität von Organismen und Organen als Gradmesser der Höherentwicklung heranzieht, kann sie nicht mehr greifen.
Strukturelle Komplexität ist eine intrinsische, dem materialen System selbst innewohnende Eigenschaft und nicht, wie etwa der Ökonomiegrad, eine relationale Eigenschaft, die von äußeren oder inneren Milieufaktoren und Vergleichsgrößen abhängt. Komplexität ist also objektiv feststellbar, ohne daß theoretische Gewichtungen und Wertungen vorgenommen werden müssen. So ist das Zentralnervensystem eines Primaten zweifellos komplexer strukturiert und daher ohne jeden Zweifel höher entwickelt, als das Strickleiternervensystem eines Insekts. An dieser Einsicht ändert die Tatsache, daß ein Insekt, welches das Gehirn eines Primaten trüge, ökonomisch benachteiligt wäre und deshalb nicht "schlechter" angepaßt oder unvollkommener ist, nicht das Geringste.
Nach KÄMPFE läßt eine Reihe weiterer intrinsischer Größen (wie etwa der Energie- und Informationsfluß in Biosystemen) Rückschlüsse auf die Entwicklungshöhe eines Organismus zu (KÄMPFE, 1992, S. 157). Beides steht eng mit dem thermodynamischen Begriff der "Entropie" (als Grad der Unordnung eines Systems) in Zusammenhang. Man kann tendenziell feststellen: Je größer der (relativ zum Gewicht gemessene) Energiefluß in einem Biosystem oder Organ, desto geringer ist seine Entropie und desto "höher" seine evolutionäre Entwicklungsstufe.
Die evolutionäre Tendenz zur Höherentwicklung und Organisation ("Entropiesenke") wird gelegentlich als Verstoß gegen den Zweiten Hauptsatz der Thermodynamik gewertet (BECK, 1978; BLISS et al. 1994). Er besagt, daß die Entropie (und damit der Grad der Unordnung) in einem System einem Maximum zustreben muß, so daß man annehmen könnte, die Entstehung geordneter Systeme sei unter natürlichen Bedingungen unmöglich.
Das Gesetz der Zunahme der Entropie gilt aber nur für adiabatisch abgeschlossene Systeme, nicht jedoch für offene Zweigsysteme fernab des thermodynamischen Gleichgewichts (MAHNER, 1986). In einem energiedurchflossenen Zweigsystem (z. B. Biosystem) können durchaus Prozesse ablaufen, die mit einer Erniedrigung der Entropie einhergehen. Den "Preis" dafür zahlt das Obersystem, dem gegenüber das Zweigsystem offen ist (KANITSCHEIDER, 1981). Das Argument ist also falsch, der Zweite Hauptsatz der Thermodynamik oft derart unverstanden, daß selbst fachlich versierte Evolutionsgegner, wie beispielsweise JUNKER und SCHERER, davor warnen, es gegen Evolution einzusetzen.
Wenn man etwa in der Wüste den Schriftzug "Coca Cola" im Sand findet, hat dessen Entropie abgenommen, die Ordnung im Sand ist gestiegen. Die Entropie im Organismus des Urhebers, gegenüber dem die Wüste "offen" ist, hat jedoch um einen noch größeren Betrag zugenommen. Auf diese Weise kann man verstehen, warum unsere Nieren - entgegen dem Konzentrationsgefälle - Stoffe aufkonzentrieren können, weshalb Pflanzen aus Kohlendioxid und Wasser Glucose aufbauen und chemische Reaktionen (wie etwa die Ammoniaksynthese) ablaufen können, obwohl die Produkte gegenüber den Edukten Entropie verloren haben. Im Falle der Biosynthese und Bioevolution bezahlt die Sonne den Preis, weil sie mehr Entropie emittiert als das Leben "abbaut".
__________________________________________
Fußnoten:
(1) Evolutionsgegner schätzen den Status fossiler Mosaikformen in aller Regel falsch ein und erkennen solche Zwischenglieder nicht als evolutionäre Übergangsformen an. Welche Argumente dahinterstehen und weshalb sie auf einem antiquierten Evolutionsverständnis beruhen, haben wir in Kapitel II.2.2 erörtert.
(2) JUNKER wendet gegen dieses Argument ein, daß es auch der Evolutionsbiologie kaum besser ergeht, weil sie nur selten Detail-Erklärungen liefern kann. Auch hier laufe die Erklärung häufig auf bloße Mutmaßungen hinaus (JUNKER, 2002, S. 74, S. 83). Trotzdem läßt sich die Situation der Evolutionstheorie mit derjenigen der Schöpfungstheorie nicht vergleichen: Wenn in der Evolutionsbiologie zahlreiche Fragen unbeantwortet geblieben sind, bedeutet das nicht, daß wir die Antworten prinzipiell nie finden können, sondern eben nur, daß wir vor praktischen Problemen der Erkenntnis stehen. Ein Biologe kann auf naturalistischer Grundlage weiterforschen, und es besteht die berechtigte Hoffnung, daß wir im Laufe der Zeit immer mehr offene Fragen beantworten können. (Man braucht sich nur einmal die Situation, in der sich DARWIN befand, mit der heutigen zu vergleichen.) Dahingegen bringt die Forschung die Schöpfungstheorie nicht weiter, weil wir prinzipiell nicht wissen können, was sich der "Designer" wohl gedacht haben mag. Insofern ist der Begriff "Schöpfungsforschung" in jedem Fall eine Zusammenstellung widersprüchlicher Begriffe.
(3) Zum einen gibt es Merkmale, die mit einem hohen Grade der Bestimmtheit als Homologien angesehen werden können. Die Einordnung fällt insbesondere bei sehr komplexen Strukturen, die teilweise bis in den mikroskopischen Bereich hinein detailgetreu bei einer großen Zahl der unterschiedlichsten Gruppen vorkommen und eventuell verschiedene Funktionen übernehmen (kurz: bei Merkmalen, deren Parallelentwicklung aufgrund von "inneren Konstruktionszwängen" nicht überzeugend erklärt werden kann), recht leicht (GOULD, 1987, S. 265). Dazu zählen, um nur einige Beispiele zu nennen, die Wirbelsäule, die Extremitäten der Tetrapoden, die Embryonalhülle, das "Dryopithecinenmuster" sowie die meisten DNA-Sequenzen vergleichbarer Proteine (Widerspruch gibt es fast nur seitens der "kritischen Schule" um GUTMANN und BONIK, auf die sich die Evolutionsgegner verschiedentlich berufen). Insbesondere die Homologie von Kontrollgenen scheint sich sich vielfach bestätigt zu haben. Wenn ein artfremder "Hauptschalter" im Organismus denselben Effekt hervorruft wie das arteigene Pendant, ist es kaum verständlich, wie sie konvergent entstanden sein sollen (GEHRING, 1998). Außerdem ist es wenig überzeugend, wenn Konvergenzen oft als unerklärte, häufig auch nicht auf Umweltanpassung rückführbare, Anomalien vorgeführt werden (vgl. z.B. JUNKER, 2002, S. 89 f.). In neuerer Zeit wird klarer, daß auch die "innere Selektion" nur bestimmte Modifikationen zuläßt, daß also das "epigenetische System" dem Gestaltungsspielraum enge Grenzen steckt. Evolution verläuft dann streckenweise in "vorgegebenen Bahnen", und gleichartige "Entwicklungszwänge" führen bei artverschiedenen Organismen zu Parallelentwicklungen, die nicht auf Abstammung und Umweltanpassung beruhen (RIEDL und KRALL, 1994, S. 263).
Copyright by Martin Neukamm, 30.08.2000
Alle Rechte vorbehalten.
Zweite, völlig neu bearbeitete Fassung (c) 02.02.2003 Last update: 30.10.2003