Allgemeines
FAQ
Publikationen
Partner
|
Vorheriger Abschnitt Zum Anfang
Druck-Version
Schritte zum Leben
Moderne Erkenntnisse über die Entstehung des Lebens
8. Erstes Leben, DNS und die Rolle der Enzyme
Der Schritt von unbelebter zu belebter Materie war also erst dann vollzogen, nachdem in den Protobionten nicht nur Stoffwechselprozesse abliefen, sondern als sich diese Zellen auch informationsgesteuert zu reproduzieren begannen. Die Codierung der Erbinformation erfolgt über Nucleinsäuren (genauer: mithilfe der Desoxyribonukleinsäure, DNS).
Die Entwicklung der ersten Nucleinsäuren dürfte viele Jahrmillionen gedauert haben. Da die DNS nicht ohne Enzyme redupliziert werden kann und Enzyme biotisch nicht ohne die DNS hergestellt werden, drängt sich die Frage auf, wie diese enge Abhängigkeit zwischen DNS und Enzymen in den ersten Lebewesen entstanden sein mag.
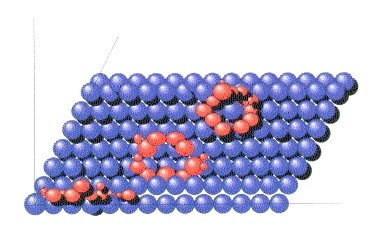
Abbildung 9:
Schematisches Modell einer rastertunnelmikroskopischen Aufnahme. Der Physiker Heckl entdeckte auf der Oberfläche von Molybdändisulfid eigentümliche ringartige und auch längliche Strukturen, deren Elektronendichteverteilung sich deutlich vom Hintergrund abhob.
Eine mögliche Erklärung liefern die Entdeckungen des Physikers Wolfgang Heckl von der IBM-Forschungsgruppe der Universität München. Heckl untersuchte diverse Trägermaterialien mit Halbleitereigenschaften auf deren Eignung zur Rastertunnelmikroskopie. Das Prinzip ist folgendes: Bringt man eine Nadel, so dünn wie ein Atom, auf eine sehr glatte leitende oder halbleitende Oberfläche, so beginnt ein Strom zu fließen, wenn man zwischen Nadel und Oberfläche eine Spannung anlegt. Die Größe des Tunnelstroms ist abhängig von der Entfernung zur Oberfläche. Dadurch lassen sich die "Täler" und "Berge" einzelner Atomlagen der Trägeroberfläche sichtbar machen.
Heckl untersuchte neben diversen synthetischen Halbleitern auch Kristalle aus Molybdändisulfid, welches wie Pyrit ebenfalls leitende Eigenschaften ausweist. Als er auf dem Bildschirm seines Computers die Atomlagen des Sulfids betrachtete, bemerkte er eigentümliche molekulare Ringstrukturen mit ca. 4 Nanometern Durchmesser sowie längliche Gebilde, die in das Trägermaterial interkaliert zu sein schienen. Die molekularen Gebilde hoben sich von der Ebene des Trägermaterials ab und waren den Strukturen in Abbildung 9 ähnlich.
Das Untersuchungsmaterial Heckl's stammte aus dem Präkambrium, war also über 650 Millionen Jahre alt. Bei näherer Untersuchung stellte sich heraus, daß diese Ringe das Element Kohlenstoff enthielten, sie mußten also organischer Natur sein. Heckl spekuliert, daß es sich hierbei um sogenannte Plasmidringe handeln könnte, also um genetisches Material sehr einfach gebauter Mikroorganismen. Diese Plasmidringe werden durch relativ kurze DNS-Sequenzen repräsentiert, die auf den Mineralien hafteten. Die Entdeckung legt den Schluß nahe: Die evolutive Erfindung des genetischen Codes sowie die enge Bindung zwischen Enzymen (die aus Aminosäuren bestehen) und der DNS (Nucleinsäure) könnte auf der Oberfläche derartiger Metallsulfide gemacht worden sein. Es ist mit Heckl denkbar, daß in der Erdfrühzeit Aminosäuren und Nucleinsäuren in mehreren Schichten übereinander auf Metallsulfiden adsorbiert wurden und so die ersten t-RNS- Stränge entstanden, die beim "Lesen" und "Übersetzen" des genetischen Codes in Proteine (bzw. Enzyme) beteiligt sind.
9. Quasispecies und Hypercyclen
Eine ganz andere Theorie verfolgt Prof. Manfred Eigen vom Max-Planck-Institut für biophysikalische Chemie in Göttigen. Seiner Ansicht nach entwickelte sich eine bestimmte Form von DNS, ein RNS-Strang, der in der Lage ist, sich ohne Hilfe von Enzymen selbst zu replizieren.
.
Abbildung 10:
Ausschnitt aus einem DNS-Molekül (Kalottenmodell). In der DNS sind zwei Fadenmolekülen spiralförmig umeinander gewickelt, die durch die Nucleotidbasen zusammengehalten werden. Die blauen bzw. organgefarbenen senkrechten Balken stellen hier die Nucleotidbasen dar; sie repräsentieren den genetischen Code. Die Spirale besteht aus einem Zucker (Desoxiribose), der chemisch mit Phosphorsäure verknüpft ist. Es existieren 4 verschiedene Nucleotidbasen: Adenin, Cytosin, Guanin und Thymin (bei der RNS anstelle Thymin: Uracil). Die Reihenfolge (Kombination) dieser Basen in dieser Doppelspirale ("Doppelhelix") charakterisiert das Genom des vorliegenden Organismus und repräsentiert dessen gesamte Erbanlagen und damit die Eigenschaften des Lebewesens. Die DNS (hier: RNS) stellt damit ein fundamental wichtiges Biomolekül dar. Beim "Lesen" der DNS wird der Doppelstrang in Einzelstränge "aufgedrillt" und die Basensequenz sukkzessive biochemisch entschlüsselt. In ihr steht "geschrieben", welche Proteine und Enzyme der Organismus herstellen muß, um Stoffwechselprozesse zu ermöglichen und .
Der "Hyperzyklus" wurde von EIGEN als erstes evolutionsfähiges Replikationssystem postuliert. In dem einfachsten Hyperzyklus finden zwei RNA-Moleküle zusammen, die sich in gegenseitiger Wechselwirkung aus einer Substratlösung hervorbringen und "vermehren". Dabei koppeln sich entweder zwei oder mehrere selbstreproduzierende RNA-Stränge (Ribozyme) oder aber Ribozyme und Enzyme zu einem stabilen Autozyklus, der sich selbst unerhält und repliziert (vgl. Abbildung 11).
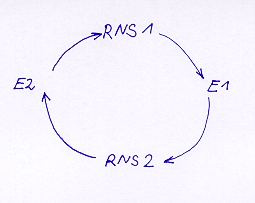
Abbildung 11:
Hier wird das schematische Prinzip eines sogenannten Hyperzyklus verdeutlicht, in dem zwei oder mehrere RNA- (RNS-) Sequenzen mit Enzymen (E) einen Zyklus bilden und sich gegenseitig erhalten. Beide dargestellten Sequenzen sind aufeinander angewiesen und können es sich nicht leisten, sich gegenseitig aus dem Rennen zu werfen.
EIGEN hatte das Modell vom Hyperzyclus ersonnen, weil die großen Fehlerraten bei der Vermehrung selbstreproduzierender Polynucleotide (Ribozyme) die Information ab einer bestimmten Sequenzlänge würde auseinanderdriften lassen. Nach jedem Replikationsschritt können nämlich Mutationen auftreten, so daß aus einer Ursequenz ein "Kometenschweif" ähnlicher Ribozyme entstehen kann. Diese bilden ein Mutantenensemble, die sogenannte "Quasispezies", die hinsichtlich Kopiergenauigkeit, Stabilität und Replikationsgeschwindigkeit miteinander in Konkurrenz stehen. Für Polynucleotide, die ausschließlich aus stabilen Guanin-Cytosin-Basenpaarungen bestehen, beträgt die experimentell bestimmte Fehlerquote etwa 1%, die Kette dürfte maximal 100 Nucleotide lang sein. Für Polynucleotide, die ausschließlich aus Adenin-Uridin-Basenpaaren bestehen, beträgt die Ablesefehlerquote bei der Replikation etwa das zehnfache, die Kettenlänge könnte also nur maximal 10 Glieder betragen. Um nun eine stabile Reproduktion ohne Informationsauflösung über beliebig viele Generationen hinweg zu gewährleisten, ist es unmöglich, ein "Ur-Genom" auf einem einzelnen Molekül zu konzentrieren.
Bilden sich in einer Quasispezies jedoch zwei oder mehrere Mutanten heraus, die ihre (eventuell durch Enzyme vermittelte) Reproduktion gegenseitig katalysieren und stabilisieren, entstehen kooperative Systeme, die sich über lange Zeiten stabil reproduzieren könnten und gegenüber allen anderen Konkurrenten im Mutantenensemble einen entscheidenden Selektionsvorteil besitzen. Als Bedingung muß gelten, daß jedes Ribozym aus 50-100 Kettengliedern besteht, für das "Ur-Gen" muß man also einen Guanin-Cytosin-Reichtum von 50-100 % annehmen, da die RNA-Matrizen lediglich dann eine hinreichend kleine Fehlerquote besitzen. Tatsächlich haben EIGEN und WINKLER-OSWATITSCH auf mathematischem Wege zeigen können, daß die rezenten t-RNA-Moleküle einen Urzustand nahelegen, der genau einer Quasispezies-Verteilung aus sich individuell reproduzierenden Molekülen entsprach (EIGEN und WINKLER-OSWATITSCH 1981). Anhand der variablen Sequenzen konnte die hypothetische Ursequenz mathematisch rekonstruiert werden (KÄMPFE 1992, S. 201).
Interessant ist ferner, daß die rezenten t-RNAs genau die angesprochenen Eigenschaften (einen hohen Guanin-Cytosin-Anteil von ca. 80% sowie eine durchschnittliche Kettenlänge von 76 Nucleotiden) und damit dieselben Zahlenverhältnisse aufweisen, die die Theorie mathematisch erwarten läßt. Nimmt man hingegen eine geheimnisvolle "creatio ex nihilo" an, bleiben die Befunde unerklärt.
10. Geschlossene Konzepte zur Entstehung der ersten Urorganismen (Protobionten)
Die ersten ganzheitlichen Ansätze zur Entstehung des Lebens auf der Basis von Nucleinsäuren wurden 1972 von Kuhn und Kaplan ausgearbeitet und bis heute stetig konkretisiert. Nach allem, was wir heute wissen, kommen zwei realistische Hypothesen in Betracht, Kuhns "Vielschritt-Hypothese" und Kaplans "Mehrtreffer-Hypothese". Im Gegensatz zu den Nucleinsäuretheorien Kuhns, Kaplans und Eigens, die postulieren, daß Oligo- bzw. Polynucleotide die primordialen Makromoleküle auf der Erde darstellten, welche ohne Replikasen bzw. Enzyme entstanden sein sollen, gehen die Proteinhypothesen (Oparin) davon aus, daß zuerst Polypeptide und Proteine entstanden, die später auf katalytischem Wege Nucleinsäuren entstehen ließen. Im folgenden wollen wir die Nucleinsäurethorien etwas eingehender erörtern und versuchen, sie mit den Proteinhypothesen und Eigens Theorie des Hyperzyclus zu verknüpfen:
a.) Vielschritt-Hypothese:
Auf Kuhns Theorie basiert die Vermutung, daß die Oligonucleotid-Sequenzen und Nucleinsäuren als primordiale Informationsträger anzusehen sind. Dabei ist wiederum die Proteinhypothese geeignet, um die Bildung dieser Nucleinsäuren, zu erklären; für diese Auffassung sprechen eine Reihe von Befunden:
Kaplan hat berechnet, daß die Wahrscheinlichkeit, daß ein Protein(oid) eine Reaktion katalysieren kann, zwischen 10-10 und 10-14 liegt. Auch die in ihrem Aufbau recht uneinheitlichen Funktionsproteinoide zeigen enzymartige Kinetiken, und man hat bereits zahlreiche derartige Proteinoide nachgewiesen. So sind abiotische Proteinoide u. a. mit ATPase-, transaminase-, esterase-, katalase- und peroxidase-Aktivitäten bekannt und es sei auf den erstaunlichen Befund hingewiesen, daß thermische Proteinoide bei Zugabe einer ATP-Lösung Oligonucleotide des Adenins aufbauen! In wässriger Lösung bei 90 °C erfolgt die Polykondensation von TMP und dAMP unter der katalytischen Wirkung von Histidin oder z. B. des Polypeptids Polyornithin, wie Oro und Mitarbeiter gezeigt haben. An derartige Oligonucleotide gelang es, stufenweise Nucleotide anzukondensieren, so daß schließlich komplexere Nucleinsäuren entstanden. Selbst die matrizenfreie, enzymgesteuerte Synthese von Polynucleotiden ist bekannt und am Beispiel der Qß-Replikase eindrucksvoll aufgezeigt worden.
Diese und andere Befunde stellen eine empirische Stütze dar, die zur Annahme berechtigt, daß sowohl Enzyme, als auch bereits einfache Funktionsproteinoide ("Protoenzyme") als (zum Teil auch matrizenfreie) Synthetasen wirken können. Die experimentellen Befunde stützen darüber hinaus Kaplans Abschätzung über die Funktionsprotein(oid)-Wahrscheinlichkeit voll und ganz. Zwar ist die katalytische Aktivität weitaus niedriger als die biotischer Enzyme, doch sie hätte zweifelsohne ausgereicht, um einfaches Leben zu ermöglichen, da "leistungsfähigere" Konkurrenten zu Beginn der biotischen Evolution noch fehlten.
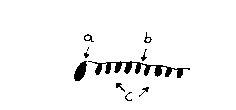
Abbildung 12:
Nucleationsmodell (a) mit Sammlerstrang (b) und Anbaumolekülen (c). Nach der Theorie konnten statistisch geknäuelte Aggregate aus Doppelhelices (a), die bestimmte Eigenschaften aufweisen, lineare offene Oligonucleotide, sogenannte Sammlerstränge binden, an deren Nucleotidbasen wiederum kurzkettige Oligonucleotide (ganz allgemein: Anbaumoleküle) über Wasserstoffbrücken gebunden wurden, die zur Entstehung der Ur-t-RNS führte.
Einfache Oligonucleotide konnten also möglicherweise auch ohne Enzyme in großer Zahl entstehen, die sich zu hyperzyklisch organisierten Quasispezies zusammenschlossen. Eine derartige Bildung erfolgte zunächst noch zufällig (divergent), mußte allerdings, wie Kuhn zeigen konnte, im Laufe der Zeit in einen konvergenten Prozeß umschlagen, der in einer Darwinschen Selektion mündete.
Dazu nehmen wir an, es bildeten sich per Zufall Oligonucleotide, deren Ribose-Zucker allesamt aus lediglich einer optische Form (etwa aus der rechtsdrehenden D-Form) bestanden. (Bei Oligonucleotiden aus z. B. 21 Gliedern ist unter 1 Million statistisch mindestens einmal eine solche Sequenz zu erwarten). Solche "enantiomerenreine" Nucleinsäuren können, wie gezeigt wurde, sehr rasch weitere, zum RNS-Strang komplementäre Nucleotidbasen binden, und recht stabile Doppelhelices aufbauen. Durch Milieuschwankungen konnten diese dann (auch das ist empirisch belegt; vgl. Kämpfe, 1992) im Zuge der konstanten Temperaturzyklen am warmen Tag wieder in Einzelstränge aufspalten, die in den kühlen Nächten mit DNS-Bausteinen derselben optischen Form wieder zu Doppelhelices rekombinierten usw. Durch derartiger Prozesse (als geeignet wird heute auch die heiße Umgebung der tiefmarinen "black smokers" angesehen), wurde also die replikasenfreie Reproduktion (Verdopplung) im Tagesrhythmus ermöglicht, wobei sich die RNS-Kopien zu Quasispezies oder tertiären Strukturen zusammenschließen konnten.Diese treten nun untereinander in einen Selektionswettbewerb, wobei Replikationsgeschwindigkeit, Kopiergenauigkeit und eventuell bestimmte katalytische, thermodynamische und sterische Eigenschaften als selektive Faktoren wirkten. Wir haben somit nach der divergenten Phase der Organisation erstmals eine echte konvergente Darwinsche Evolution vorliegen; Selektion führt automatisch zu Informationsaufbau und Negentropie. Auch die optische Symmetrie der Riboseeinheiten läßt sich mit diesem Modell ganz zwangsläufig erklären, da nur uniforme, enantiomerenreine RNS-Oligomere die Bildung stabiler, kopiergenauer und hochreplikativer Doppelhelices gewährleisten können.

Abbildung 13:
Auf einer Aggregat-Oberfläche (a) (Sammlerstrang) können Anbaumoleküle (b) (z. B. kurze offenkettige Oligonucleotide) gebunden werden, die ihrerseits eine starke Affinität zu Aminosäuren (c) aufweisen. Das gesamte Aggregat kann, wie Wächtershäuser glaubt (siehe Theorie des Biofilms, Oberflächenmetabolismus) auf der Oberfläche von Pyritkriställchen, Tonen, Sand oder Lava haften bzw. entstehen, wobei Pyrit und andere Metallsulfide über günstige katalytische Eigenschaften, etwa auch zur Entstehung von Proteinen besitzen (vgl. empirische Befunde von Cairns-Smith et. al.), die die Verknüpfung der Aminosäuren zu einem primärstrukturierten Proteinstrang bewerkstelligten; der ursprüngliche Übersetzungsapparat war entstanden. Im Laufe der Zeit entstanden dann evolutiv geeignete Enzym-Nucleinsäure-Hypercyclen, die ein selbstreplizierendes, autokatalytisches System bildeten.
Nun weiß man, daß sich Nucleinsäuren mit Tertiärstruktur zu statistischen Knäueln verbinden können, wobei derartige Aggregate unter bestimmten Bedingungen wiederum zahlreiche nadelförmige Oligonucleotide zu binden in der Lage sind, die wie "Stecknadeln aus einem Nadelkissen" herausragen. Diese geradlinigen Oligonucleotide bezeichnete Kuhn als "Sammlerstränge", die, wie es unsere heutigen Kenntnisse nahelegen, wohl in der Lage waren, ihrerseits wieder kurze komplementäre Polynucleotid-Sequenzen ("Anbaumoleküle") über Wasserstoffbrücken zu binden. Die Anbaumoleküle bildeten nach Kuhn die Ur-t-RNAs, die Sammlerstränge die Ur-m-RNAs. Diese recht komplexen Aggregate können Aminosäuren an den Anbaumolekülen anlagern, die Bildung von Proteinen und komplexen Enmzymen ist dann relativ wahrscheinlich. Neben den Aggregaten kommen nach Katchalsky aber auch anorganische Tracersubstanzen mit katalytischen Eigenschaften in Frage, wie Pyrit oder Montmorillionit. Wie aus Abbildung 12 und 13 ersichtlich ist, bildete sich der ursprüngliche Übersetzungsapparat dadurch heraus, daß die Aminosäuren an die Anbaumoleküle gebunden und durch die Katalyse der Aggregatoberflächen zu Enzymen umgesetzt wurden. Der Brückenschlag zu Wächtershäusers Theorie des Oberflächenmetabolismus (vgl. oben) ist augenfällig.
b.) Mehrtreffer-Hypothese
Im Gegensatz zu Kuhns Vielschritt-Hypothese postulierte Kaplan eine Entstehung der ersten Protobionten in einem Schritt, wobei zufällig Synthetasen und Nucleinsäuren in einem Kompartiment zusammengefunden haben mußten. Von jeder der beiden Funktionseinheiten mußte ein "Basisrepertoire" an Makromolekülen vorhanden gewesen sein, die sich selbst zu autokatalytischen Verbänden organisierten. Kaplan schätzte die Rahmenbedingungen ab und kam nach statistischen Überlegungen zum Schluß, daß die Bildung derartiger selbstreplikativer Systeme durchaus im Bereich des Wahrscheinlichen gelegen haben könnte (vgl. Kämpfe, 1992).
Holleman, Wiberg: Lehrbuch der Anorganischen Chemie, S. 518 f. Berlin, 1995
E. W. Bauer et al: Biologiekolleg. Bielefeld, 1983
Bilder frühen Lebens. Verständliche Forschung, Spektrum der Wissenschaft. Heidelberg, 1986
Entwicklung von den ersten Lebensspuren bis z. Menschen. Verständl. Forschung, Spektrum d. Wiss.. Heidelberg, 1988
Darwin, Charles: Die Entstehung der Arten durch natürliche Zuchtwahl. Stuttgart, 1963
Hoimar v. Ditfurth: Am Anfang war der Wasserstoff. Hoffman und Campe, 1980
Stuart Kauffman: Der Öltropfen im Wasser, 1995
L. Kämpfe: Evolution und Stammesgeschichte der Organismen, Gustav-Fischer-Verlag, 1992
Last update: 07.05.05
Vorheriger Abschnitt Zum Anfang © M. Neukamm , 22.06.99